Предлагаемая читателю работа депонирована в ВИНИТИ РАН, и ниже перед титульным листом полностью приведены её выходные данные и текст автореферата, предназначенного для публикации в Реферативном журнале ВИНИТИ.
С автором, Львом Николаевичем Солнцевым, можно связаться по телефону 335-64-20.
УДК 595.78
Выведение бабочек (Lepidoptera) без диапаузы./ Л.Н.Солнцев; ВНИИ охраны природы. М., 2002. - 15 с. - Библиогр. 5 назв. - Рус. (Рукопись деп. в ВИНИТИ 22.04.2002, N 731-В2002)
В лабораторных условиях без диапаузы при повышенной температу- ре (20 - 22oC) успешно выведены 54 вида из семейств Lasiocampidae, Arctiidae, Lymantriidae, Noctuidae, Geometridae, Saturniidae. Способность диапаузировать или под влиянием изменения внешней среды развиваться без диапаузы обусловлена генотипически, зависит от видовой и родовой принадлежности бабочек.
При скрещивании и получении нескольких "бездиапаузных" поколений Spilosoma niveum и Arctia caja скорость развития от яйца до имаго увеличивается в несколько раз (пример быстрых неканонических наследственных изменений). Приведены практические советы по бездиапаузному воспитанию бабочек.
В работе изложены основные методологические принципы и практические советы бездиапаузного воспитания бабочек (Macrolepidoptera) ряда семейств - Lasiocampidae, Arctiidae, Lymantriidae, Noctuidae, Geometridae и Saturniidae. Эти положения сформулированы на основании многолетних опытов автора по выведению бабочек без диапаузы в искусственных условиях.
С уменьшением длины светового дня и приближением осени, а затем зимы, для преимагинальных стадий бабочек наступает диапауза - особое физиологическое состояние насекомого, находящегося в покое, которое позволяет выжить в неблагоприятных климатических условиях. К числу таких неблагоприятных условий относятся не только наступление морозов, но и засуха, отсутствие кормовой базы. В то же время чешуекрылые в течение полного развития от яйца до имаго подвергаются многочисленным опасностям. Наличие паразитирующих насекомых и грибковых заболеваний, а также непродуманная человеческая деятельность, трансформирующая окружающую среду, в значительной степени сокращают численность и видовой состав чешуекрылых. С учётом этих обстоятельств ускоренный бездиапаузный метод воспитания бабочек в искусственных условиях имеет ряд преимуществ и может иметь исключительное значение для восстановлений нарушенной энтомофауны и в некоторых других случаях.
В лабораторных условиях при температуре 20 - 22 градуса Цельсия и постоянной влажности для выведения бабочек создаётся стабильный микроклимат, позволяющий организовать необходимый режим питания и содержания гусениц, а также подобрать кормовое растение с учётом его фенологического состояния в сочетании с питательными пищевыми добавками.
Рациональная технология воспитания бабочек ускоряет процесс развития, в результате которого за 6 - 7 месяцев можно получить до 5 поколений хорошо окрылённых и в большом количестве особей разного пола для дальнейшего скрещивания с целью проведения опытов по изучению биологии, экологии и этологии чешуекрылых.
Разведение бабочек с последующим расселением их в природе открывает широкие перспективы восстановления энтомофауны парков и лесопарковых зон Москвы и Московской области.
Заслуживает внимания идея промышленного бездиапаузного воспроизводства бабочек, особенно, если учесть, что кладка, например, одной из самок Spilosoma niveum (Men) или Arctia caja (L.), насчитывает тысячу яиц, а при получении нескольких поколений общее количество бабочек может быть использовано в виде биомассы в различных областях народного хозяйства.
Успех воспитания гусениц зависит от родовой и видовой принадлежности бабочек, а также выбора преимагинальной стадии. Грена, оплодотворённой самки, и гусеницы в молодом возрасте являются предпочтительным материалом для эксперимента. Гусеницы после второй-третьей линьки безболезненно приспосабливаются к тепловому режиму в новой среде обитания.
В природных условиях изменение суточного режима температуры, влияние фотопериодической реакции и физиологического состояния кормовых растений служат для гусениц источником сезонной информации наступления диапаузы [Данилевский, 1961; Дружелюбова, 1980]. В этот период у гусениц изменяется биохимический состав крови в сторону увеличения соли и глицерина - веществ, увеличивающих сопротивляемость организма промерзанию. Поэтому гусеницы, собранные в сентябре месяце и приготовившиеся зимовать, с большим трудом поддаются бездиапаузному воспитанию.
Первостепенной задачей выращивания гусениц является выбор кормового растения [Мкртумян, 1958], особенно в осенне-зимний период. В условиях Москвы и Московской области такой кормовой базой послужила ива корзиночная (Salix viminalis L.), сохраняющая свою сочную листву вплоть до зимних заморозков, ряд травянистых растений с растянутыми сроками вегетации (одуванчик лекарственный - Taraxacum officinale Wigg., клевер луговой - Trifolium pratense L., тысячелистник обыкновенный - Achillea millefolium L., ежа сборная - Dactylis glomerata L., глухая крапива - Lamium album L.), а также растения, которые растут над тепловыми инженерными трассами (трубы городского горячего водоснабжения подогревают слой почвы и создают эффект парника). Биохимический состав (насыщенность сахарами и другими веществами) и высокая энергетика растений стимулируют рост гусениц, ускоряют развитие бабочек.
Без осложнений развиваются гусеницы рода Spilosoma, Callimorpha, Phragmatobia, Hyphoraia, Parasemia, Rhyparie, Arctia, Hipocrita (за исключением Arctia hebe (L.) - Штандфусс, дата издания книги не указана); гусеницы волнянок рода Dasychira, Orgyia; многие гусеницы совок рода Acronicta, Diarsia, Agrotis, Aplecta, Polia, Anarta, Blepharita.
Шелкопряды выводятся значительно хуже, чем медведицы. Опыты, проводимые в течение нескольких лет над Macrothylacia rubi (L.), показали, что для воспитания этого вида необходимо иметь 40 - 50 гусениц, чтобы получить 2 - 3 имаго преимущественно самок.
Гусеницы из грены вылупляются все одновременно. Затем одни гусеницы быстро взрослеют, окукливаются и вскоре выводятся бабочки; другие - замедляют свой рост, и бабочек из куколок выходит небольшой процент; остальная часть гусениц впадает в диапаузу.
Отмеченная закономерность особенно четко прослеживается у Arctia caya (L.). В первую очередь из гусениц одной и той же грены выводится 70 %% бабочек; во вторую, со значительным отставанием, - 20 %% и 10 %% гусениц, достигнув 1,5 - 2 см в длину, прекращают развиваться. Известно, что основой развития бабочки служит генетически обусловленная программа, в которой запрограммировано время созревания преимагинальных стадий, и воздействие внешней среды только активизирует или замедляет этот процесс, способствуя иногда образованию новых аберраций.
Задача энтомолога-экспериментатора, учитывая генотипически обусловленные закономерности развития вида, найти пути и средства, препятствующие у гусениц наступлению преддиапаузного состояния. Часть гусениц, которые перестают питаться и проявляют признаки наступающей диапаузы, иногда целесообразно, чтоб вывести их из оцепенения, искупать в тёплой воде (40 - 50 градусов Цельсия) с одновременным лёгким массажем с целью размягчить остатки растительной пищи в кишечнике. Воздействие повышенной температуры и облучение ультрафиолетовыми лучами, смена кормового растения, смоченного подслащенной водой или соком апельсина с мёдом, включение в пищевой рацион овощей (листья капусты, петрушки, укропа), фруктов (дольки мандарина, кожура яблока и банан) способствуют дальнейшему развитию гусениц.
В природе при неблагоприятных климатических условиях и отсутствии питания гусеницы некоторых видов бабочек не погибают, а начинают диапаузировать, в особых случаях, в стадии куколок - до нескольких лет. Подобное явление произошло с гусеницей бабочки Carcinopyga proserpina Standfuss, которая воспитывалась при повышенной температуре, и была вынуждена, из-за отсутствия кормового растения, пробыть в диапаузе свыше месяца.
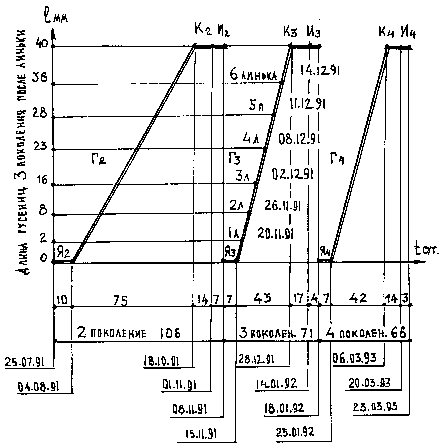
График, представленный ниже (рис. 1), иллюстрирует развитие второго, третьего и четвёртого поколения бабочки Spilosoma niveum (Men) (первое поколение не показано, поскольку оно развивалось в природных условиях). Время, в продолжении которого происходит созревание преимагинальных стадий, отложено на оси абсцисс (t в сутках). На оси ординат отмечена длина гусениц после линек, а на графике - даты, когда это произошло. Третье и четвертое поколения развивались в два раза быстрее, чем второе. Замедленный рост второго поколения можно объяснить факторами, связанными с постепенным переходом гусениц в бездиапаузное состояние (влияние температуры, новых кормовых растений и т.д.).
Внешний вид бабочки Spilosoma niveum (Men), по сравнению с экземплярами, выведенными в естественных условиях, от поколения к поколению, оставался неизменным, как по рисунку, окраске, так и по форме крыльев.
Рис. 1. График бездиапаузного развития преимагинальных стадий нескольких поколений бабочки Spilosoma niveum (Men.).
Условные обозначения к рис. 1 и 2: Я 2,3,4 - стадии развития яиц; Г 2,3,4 - стадии развития гусениц; К 2,3,4 - стадии развития куколок; И 2,3,4 - стадии развития имаго. 1 мм - длина гусениц в мм.; t сут. - время в сутках.
Опыты по многократному скрещиванию проводились на бабочках Arctia caya (L.) (рис. 2). 40 гусениц указанного вида были собраны в мае вблизи пл. Воймежный Казанской ж.д. 50 %% гусениц оказались поражёнными мухами. С конца июня по 09.07.1996 г. вывелось 20 бабочек первого поколения. 8 самок отложили яйца. Полный цикл развития второго поколения от яйца до имаго продолжался 86 суток.
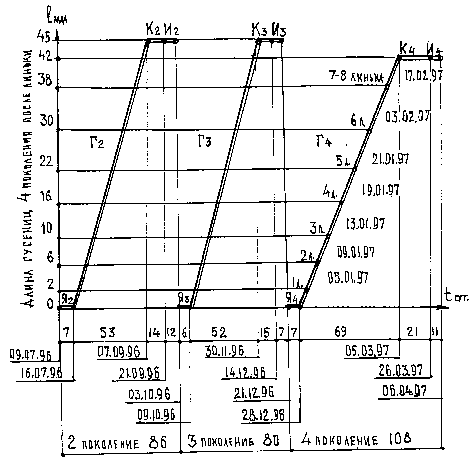
Рис. 2. График бездиапаузного развития преимагинальных стадий нескольких поколений бабочки Arctia caja (L.).
Условные обозначения - см. подпись к рис. 1.
Третье поколение развивалось несколько быстрее второго - 80 суток. Имаго третьего поколения вывелись нормально окрылёнными,обычных размеров, без изменения в окраске и рисунке крыльев.
Полный цикл развития четвёртого поколения составил 108 суток. Рост гусениц замедлился, количество линек увеличилось до 7 - 8. Гусеницы, достигнув длины 1 - 1,5 см, впали в диапаузу. Бабочки четвёртого поколения оказались меньше размером, апекс верхних крыльев закруглён, цветовая гамма поблёкла, рисунок пятен несколько изменился. На замедленное развитие четвёртого поколения, возможно, повлияло недоброкачественное кормовое растение, случайно попавшее в пищу гусеницам. Кладки вновь выведшихся самок оказались не оплодотворёнными. Были замечены случаи каннибализма, когда гусеницы поедали яйца, отложенные бабочками.
Для выведения имаго, скрещивания и кладки яиц рекомендуется садок, состоящий из деревянного ящика размером 15 x 30 x 6 см, на дно которого помещается ванночка с водой (рис. 3). Ящик покрывает картонный объём в виде усечённой четырёхгранной призмы высотой 15 см. В верхней части призмы имеется вентиляционное отверстие, закрытое сеткой, не допускающей проникновение крошечных рыжих фараоновых муравьёв (Monomorium pharaonis L.). На выступы стенок ящика устанавливается рамка с натянутой мелкой сеткой из капрона, на которую стелится, смоченная водой и отжатая, льняная тряпочка. Куколки, предварительно завёрнутые в кусочки бумажной салфетки, укладываются рядами и покрываются также влажной льняной тряпочкой. Коконы лежат свободно и только периодически сбрызгиваются водой. Стенки призмы изнутри выстилаются марлей, на которую самки бабочек кладут яйца. Кладка с кусочком марли вырезается и помещается, вместе с соответствующим кормовым растением, в обычную литровую или пол-литровую стеклянную банку. На дно банки кладётся бумажная салфетка. Сверху банка покрывается марлей в два слоя.
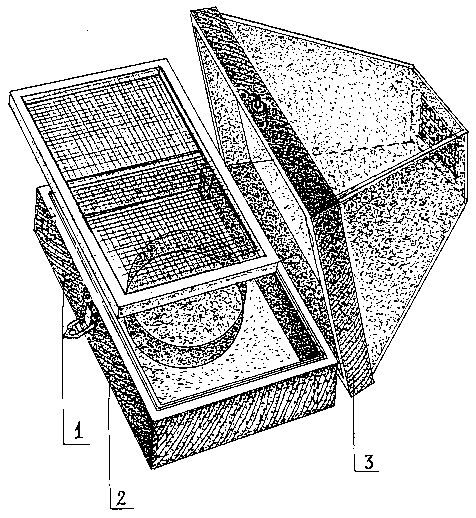
Рис. 3. Садок для выведения бабочек.
При содержании и кормлении гусениц в банках необходимо следить, чтобы не было переувлажнения от излишне положенного кормового растения. Не рекомендуется банки с гусеницами ставить под прямые лучи солнца, вызывающие отпотевание стенок. Обилие влаги и перегрев могут отрицательно сказаться на развитии только что выведшихся гусениц. Особое внимание следует уделить линьке гусениц. В период смены шкурки гусеницы перестают питаться и, прикрепившись к стенке сосуда, вступают в особое биологическое состояние, которое легко спутать с наступлением диапаузы. Гусеницы после линьки сразу же начинают жадно пить подслащенную воду и активно поедать корм.
Продолжительное изучение преимагинальных стадий чешуекрылых и многолетняя работа по воспитанию гусениц бездиапаузным методом позволило получить свыше 50 видов бабочек различных родов и семейств.
Указывается число выведенных особей каждого вида, из чего они выведены (из гусеницы или из яйца), дата и место поимки исходных экземпляров.
Eudia pavonia (L.), 3 экз., e.l., 27.10.1985; п. Салтыковка Московской области. III тип.
Macrothylacia rubi (L.), 2 экз., 25.10.1982; 2 экз., e.l., 21.11.1986; п. Фосфоритный Московской области. III тип.
Calliteara pudibunda (L.), 2 экз. e.l., 17.11.1988; там же. II - III типы.
Gynaephora selenitica (Esp.), 9 экз., e.l., 16.11.1985, Калуга-2. III тип (в основном, выводятся самки).
Euproctis simmilis (Fuessly), f. auriflua (Hbn.), 5 экз., e.l., 28.9.1991, Приморский край, г. Владивосток. II тип.
Spilarctica cosigneta (Koll.), f. seriatopunctata Motsch., 7 экз., e.l., 17.9.1991; Приморский край, г. Владивосток. II тип.
Hyphorai aulica (L.), 8 экз., e.l., 5.11.1985; п. Фосфоритный Московской области. II тип.
Diacrisia sannio (L.), 1 экз., e.l., 13.11. 1986, там же. II тип.
Rhyparia purpurata (L.), 8 экз., e.l., 11.10.1989, там же. II тип.
Parasemia plantaginis (L.), 3 экз., e.l., 12.10.1991, там же. II тип.
Phragmatobia fuliginosa (L.), 4 экз., e.l., 29.10.1989, там же. II тип.
Spilosoma lubricipedum (L.), 2 экз., е.l., 02.11.1992, п. Воймежный Московской области. II тип.
Spilosoma niveum (Men.), 3 поколения, е.о., с 25.07.1991 по 23.02.1992, Приморский край, г. Владивосток. I тип.
Callimorpha dominula (L.), 3 экз., е.l., 04.01.1995, п. Дорохово Московской области. II тип.
Carcinopyga proserpina Standfuss, 3 экз., е.о., 28.02.1981, Узбекистан, Ташкент. (03.10.1980 вывелись гусеницы). III тип.
Arctia caja (L.), 3 поколения, е.о. с 09.07.1996 по 06.04.1997, п. Воймежный Московской области. II тип.
Agrotis exclamationis (L.), 1 экз., e.l., 29.11.1992, п. Фосфоритный Московской области.
Xestia c-nigrum (L.), 1 экз., е.l, 31.10.1989, там же.
Xestia ditrapesium (Den. et Schiff.), 1 экз. е.l., 10.11.1988, там же.
Diarsia brunnea (Den. et Schiff.), 1 экз. е.l., 10.11.1988, там же.
Diarsia rubi (View.), 1 экз., e.l., 27.10.1985, там же.
Paradiarsia punicea (Hbn.), 8 экз., e.l., 01.11.1985, там же.
Eurois occulta (L.), 5 экз., е.l., 16.10.1987, там же.
Mamestra brassicae (L.), 1 экз., е.l., 11.10.1995, п. Фосфоритный Московской области.
Lacanobia contiqua (Den. et Schiff.), 2 экз., е.l., 20.10.1985, там же.
Lacanobia thalassina (Hfn.), 1 экз., е.l., 17.11.1989, там же.
Lacanobia pisi (L.), 1 экз., е.l., 29.11.1996, там же.
Lacanobia w-latinum (Hfn.), 1 экз., e.l., 09.11.1994, там же.
Melanchra persicariae (L.), 1 экз., e.l., 15.11.1994, там же.
Discestra trifolii (Hfn.), 2 экз., e.l., 08.10.1987, там же.
Polia hepatica (Cl.), 1 экз., e.l., 11.11.1994, там же.
Polia bombicina (Hfn.), 6 экз., e.l., 12.11.1990, там же.
Polia nebulesa (Hfn.), 4 экз., e.l., 17.11.1985, там же.
Anarta myrtilli (L.), 5 экз., e.l., 21.10.1984 - 1994, там же.
Aletia pudorina (Den. et Schiff.), 3 экз., e.l., 28.11.1989, там же.
Euplexia lucipara (L.), 1 экз., e.l., 20.11.1994, там же.
Mesapamea secalis (L.), 1 экз., e.l., 23.11.1989, там же.
Enargia paleacea (Esp.), 11 экз., e.l., 11.10.1992, там же.
Dicycla oo (L.), экз., e.l., 15.10.1990, там же.
Athetis palustris (Hbn), 2 экз., e.l., 15.11.1987, там же.
Apamea crenata (Hfn.), 3 экз., e.l., 17.11.1985, там же.
Apamea sordens (Hfn.), 1 экз., e.l., 30.10.1987, там же.
Acronicta auricoma (Den. et Schiff.), 2 экз., e.l., 06.11.1987, там же.
Acronicta rumicia (L.), 10 экз., e.l., 27.11.1990, там же.
Acronicta euphorbiae (Den. et Schiff.), 2 экз., e.l., 18.11.1989, п. Воймежный Московской области.
Acronicta menyanthidis (View.), 1 экз., e.l., 12.11.1987, п. Фосфоритный Московской области. Вывелся наездник.
Acronicta megacephala (Den. et Schiff.), 1 экз., e.l., 07.11.1987, п. Воймежный Московской области.
Acronicta tridens (Den. et Schiff.), 1 экз., e.l., 12.11.1990, п. Фосфоритный Московской области.
Acronicta psi (L.), 12 экз., e.l., 06.11.1987, там же.
Blepharita adusta (Esp.), 2 экз., e.l., 24.11.1990, там же.
Blepharita bathensis Lutzau, 3 экз., e.l., 29.11.1986, там же.
Angerona prunaria (L.), 1 экз., e.l., 08.11.1988, п. Фосфоритный Московской области. II - III типы.
Semiothisa clathrata (Cl.), 3 экз., e.l., 28.10.1987, там же. II - III типы.
Pelarga comitata (L.), 1 экз., e.l., 23.10.1987, там же. II - III типы.
Способность бабочек диапаузировать или развиваться без диапаузы под влиянием изменения внешней среды обусловлена генотипически.
Бездиапаузное развитие гусениц зависит от видовой и родовой принадлежности бабочек.
При скрещивании и получении нескольких "бездиапаузных" поколений, скорость развития преимагинальных стадий от яйца до имаго увеличивается в несколько раз.
Данилевский А.С. Фотопериодизм и сезонное развитие насекомых. Л., Изд-во Лен. гос. ун-та, 1961. 241 с.
Данилевский А.С. Роль питающих растений в биологии лугового мотылька. - Энтомологическое обозрение. Л., 1936. Т. 26, вып. 1 - 4. С. 91 - 110.
Дружелюбова Т.С. Влияние температурного и светового факторов на развитие и морфофизиологические особенности генерации лугового мотылька. - Тр. ВИЗР. Эколого-физиологические предпосылки современной системы борьбы с луговым мотыльком. Л., 1980. С. 69 - 81.
Мкртумян К.Л. Роль различных кормовых растений в развитии мальвовой моли в Армянской ССР. - Тр. Ин-та земледелия. Ереван, 1958. Вып. 1. С. 69 - 79.
Штандфусс М. Жизнь бабочек, их ловля, воспитание и сохранение. Руководство для собирателей. СПб, Невский пр-кт 50, [дата издания книги на ней не указана]. 315 с.
Материал размещён 06 апреля 2003 г.